
Qu’est-ce-que les haptènes ? > Un petit cours d’immunologie
Les groupes sanguins
C’est un médecin et biologiste autrichien, Karl Landsteiner, qui a découvert, dans les années 1900-1901, les premiers groupes sanguins. Il a pu déterminer les groupes sanguins en fonction de la présence ou de l’absence de certains antigènes sur les membranes des érythrocytes.
Il a proposé aussi d’attribuer des chiffres : I, II, III, IV pour dénommer les groupes sanguins. C’est à Ludwik Hirszfeld von Dungern qu’on doit les appellations A, B, AB, et 0 (zéro, changée plus tard en lettre « O »). Ces appellations ont été adoptées dans le monde entier en 1928.
Ludwik Hirszfeld, un médecin microbiologiste et sérologiste polonais, a été le premier à prévoir une existence possible du conflit sérologique entre la mère et l’enfant, confirmé plus tard par la découverte du facteur Rhésus. Il a aussi découvert l’hérédité des groupes sanguins.
Les érythrocytes
Appelés globules rouges, les érythrocytes sont les cellules du sang les plus nombreuses.
Dans un millilitre cube de sang, il y a 5 millions d’érythrocytes. Leur rôle principal est le transport d’oxygène depuis les poumons vers les tissus par l’intermédiaire de l’hémoglobine.
La durée de vie des érythrocytes dans l’organisme est d’environ 120 jours. Un érythrocyte est une cellule en forme de disque biconcave. Comme il est souple, il peut changer de forme en circulant dans les capillaires.
Si l’on pouvait ranger des érythrocytes à plat, côte à côte, sur la longueur d’un millimètre, on pourrait compter 133 érythrocytes. Si l’on les rangeait en forme de colonne de hauteur d’un millimètre, on y trouverait 400 érythrocytes. Le résultat du calcul donne le nombre de 7 millions d’érythrocytes environ dans un millilitre cube.
Dans un organisme sain, les érythrocytes restent en suspension dans la lymphe, séparés les uns des autres car ils sont chargés négativement. Plus une charge négative est grande et plus grande est la force qui les repousse. Une charge faible facilite l’agglutination des érythrocytes.
L’unité de mesure de la charge électrique est coulomb (C). On ne dispose pas d’appareil pour mesurer les charges électriques des érythrocytes. Mais la vitesse de sédimentation permet une estimation approximative des charges électriques.
Le phénomène de sédimentation dans un échantillon de sang non coagulé a été décrit d’une façon détaillée par un chercheur polonais, Edmund Faustyn Biernacki. Cette analyse porte son nom, c’est la réaction de Biernacki. Elle est appelée aussi la VS ou ESR. Elle consiste à mesurer la vitesse à laquelle les érythrocytes tombent au fond d’un tube.
Le résultat normal de VS est de 2 à 8 mm/h,et il signifie que les charges électriques sont suffisamment élevées. Si les valeurs sont en dessous de 8 mm/h, les charges électriques sont trop faibles.
Il est aussi possible de déterminer les charges électriques des érythrocytes en mesurant le potentiel zêta sur le voltmètre. Le potentiel zêta normal dépasse 35 mV. Le résultat : 15 – 35 mV est une stabilité moyenne ; 5 – 15 mV est la limite de stabilité et le seuil d’agglutination ; 0 – 5 mV est un potentiel favorable à l’agglutination. Si le potentiel zêta est normal, les érythrocytes gardent entre eux une distance minimum de 25 nm.
Les leucocytes
Les leucocytes ou globules blancs participent à la défense de l’organisme contre des agents pathogènes. Il en existe plusieurs types : granulocyte, lymphocytes, monocytes qui forment un système de défense bien organisé. Leurs dimensions sont comprises entre 5 000 et 20 000 nm. Un individu en bonne santé possède de 6 000 à 8 000 leucocytes dans un millilitre cube de sang. Chez un malade de leucémie, le taux de leucocytes est augmenté. Dès lors, les leucocytes perdent de l’efficacité, et ils peuvent même attaquer des cellules de l’organisme, en entraînant alors des maladies auto-immunes.
Les antigènes immunogènes
Ce sont des molécules composées de quelques milliers à quelques centaines de milliers d’atomes.
Ces grosses molécules sont le plus souvent des protéines, des polysaccharides, des lipides et des acides nucléiques. Leur masse est comprise entre 10 000 et quelques millions d’unités de masse atomique.
Les antigènes sont très diversifiés, mais tellement petits qu’on ne peut les voir ni à l’œil nu, ni à la loupe, ni au microscope optique ordinaire. Les techniques de biologie moléculaire ont dévoilé les formes, les dimensions, les masses moléculaires et les propriétés des antigènes et des différentes molécules qui composent les organismes vivants.
Les antigènes immunogènes ont la capacité de stimuler la production d’anticorps dans l’organisme, s’ils sont introduit dans cet organisme par une autre voie que le système digestif (par l’injection ou une blessure). Les antigènes introduits par voie orale sont le plus souvent digérés et détruits. Les anticorps produits par l’organisme se lient aux antigènes spécifiques, ceux qui ont provoqué la production de ces anticorps ou bien aux antigènes similaires. La liaison se fait aux endroits appelés déterminants antigéniques qui se trouvent sur les antigènes.
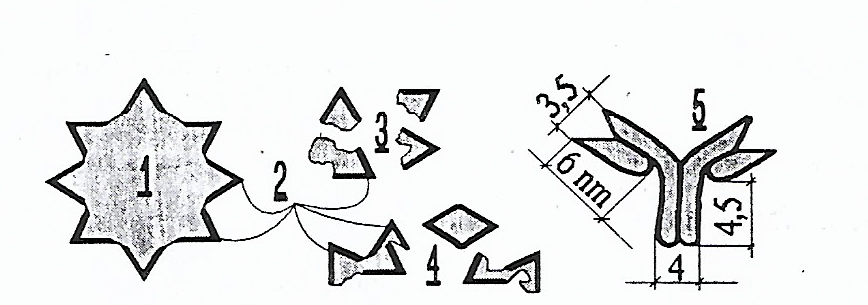
Chaque individu possède sur ses érythrocytes divers « paquets » d’antigènes. De nombreux déterminants antigéniques existent non seulement sur les membranes des érythrocytes mais aussi sur les membranes des leucocytes, des plaquettes et différentes cellules.
Les antigènes majeurs de surface sur les érythrocytes sont les antigènes A, B (du système ABO) et D (du système Rh). Mais le fait de connaître le groupe sanguin ne suffit pas pour pouvoir déterminer le genre de déterminants existant sur d’autres cellules du corps.
Les analyses d’antigènes présents sur les plaquettes, les lymphocytes et d’autres cellules ne sont pas incluses dans les analyses de routine. En général, on effectue des analyses concernant les plus importants, les plus forts antigènes immunogènes : A, B, D et H.
L’antigène H est un antigène public. Il est présent chez presque tous les humains (99,99%), mais la quantité de cet antigène n’est pas identique chez tout le monde. Il dépend du groupe sanguin, ce qu’on peut présenter par l’ordre décroissant, comme suit :
0 > A2 > B > A2B > A1 > A1B
Les anticorps
Les anticorps, appelés aussi immunoglobulines, présents dans le sang (le sérum) sont capables de se lier aux antigènes étrangers : les anticorps anti- A se lient aux antigènes A, les anticorps anti- B se lient avec les antigènes B. Les autres anticorps se lient avec les antigènes spécifiques. Les anticorps sont des protéines complexes. Ce sont de grosses molécules de masse moléculaire de 150 à 900 kD .
On distingue : les anticorps naturels et les anticorps immuns.
Selon les groupes sanguins, on peut trouver différentes variétés d’anticorps, mais leurs quantités varient en fonction de l’âge de l’individu donné.
Les anticorps naturels apparaissent dans l’organisme spontanément, au cours du troisième mois après la naissance. Avant, l’organisme du nouveau-né ne produit pas d’anticorps et ceux qu’on trouve chez lui proviennent de l’organisme de la mère. Les parois très fines du système digestif du nouveau-né allaité par sa mère laissent passer facilement de grandes quantités d’anticorps.
Durant les premières années de la vie de l’enfant, le taux d’anticorps naturels est bas, mais il augmente progressivement jusqu’à l’âge de 10 ans, et c’est à cette période que le taux d’anticorps naturels est le plus élevé. Puis, il diminue au fil du temps.
Le taux d’anticorps naturels β ( anti-B) chez des jeunes organismes avec le groupe sanguin A atteint habituellement environ 200.
Le taux d’anticorps naturels α ( anti- A) chez des jeunes organismes avec le groupe sanguin B atteint habituellement environ 350.
Le taux d’anticorps naturels chez des jeunes organismes ayant le groupe sanguin 0 : α (anti-A) atteint le plus souvent le nombre 400, et β (anti-B) : environ 170.
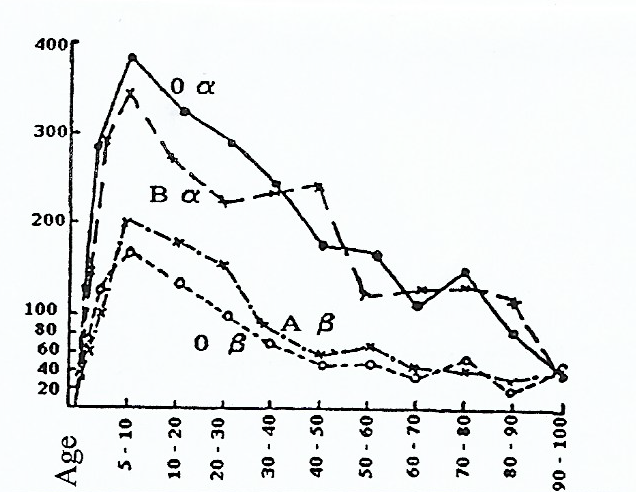
Un taux d’anticorps naturels dans le sang humain, selon Thomsen et Kettl, 1929.
Cependant, les personnes du même âge et de même groupe sanguin peuvent présenter de différents taux d’anticorps naturels.
Il existe des classes d’anticorps (immunoglobulines) selon leur masse moléculaire. Le sang humain contient : Ig G – 80%, IgA – 13% , IgM – 6%. Les autres classes sont minoritaires.
- Les immunoglobulines IgG sont les plus abondantes et les plus petites : leur masse est de 150 kD environ. Leurs petites dimensions facilitent le passage du sang maternel vers le sang du foetus à travers le placenta, composé de trois couches, ce qui peut causer parfois un conflit sérologique.
- Les immunoglobulines de classe IgA se trouvent dans le sang, mais essentiellement dans les sécrétions comme la salive, le lait maternel, la sueur, les larmes…).
- Les immunoglobuline de classe IgM sont sécrétées rapidement lors du premier contact avec un antigène.
- Les anticorps naturels anti-A appartiennent à la classe IgA et IgM.
- Les anticorps naturels anti-B appartiennent à la classe IgM.
- Les anticorps naturels anti-Lea et anti-Le b appartiennent à la classe IgM.
- Les anticorps immuns anti-A, anti-B, anti-Le a, anti-Leb et plusieurs autres anticorps appartiennent surtout à la classe IgG. Mais au début de la réponse immunitaire (primaire et secondaire), ce sont les IgM qui sont fabriquées d’abord et les IgG apparaissent plus tard. Les anticorps IgM atteignent un taux élevé en phase initiale. Leur production diminue après un cours laps de temps, et ce sont les IgG surtout qui restent dans l’organisme le plus longtemps.
Les différences entre les anticorps naturels et les anticorps immuns sont comme suit :
- Le taux d’anticorps naturels anti-A et anti-B évolue lentement et dépend de l’âge de l’individu donné ;
- Les anticorps immuns se forment après la stimulation antigénique dû à la présence d’un antigène spécifique (immunogène), et leur taux évolue de deux façons :
- Après un premier contact, le taux d’anticorps monte lentement, et il n’est pas élevé ;
- Des contacts suivants avec un même antigène provoquent au début un abaissement de courte durée suivi d’une production d’anticorps intense et rapide.
Étant donné la classification des antigènes en deux catégories : les anticorps naturels et les anticorps immuns, il serait judicieux de distinguer la sensibilisation naturelle et la sensibilisation immune.
La sensibilisation
La sensibilisation de l’organisme a lieu à la suite de l’introduction de micro-organismes ou de substances immunogènes, et elle est conditionnée par :
- infections virales, infections bactériennes, parasitoses ;
- transfusion de sang provenant d’un donneur ;
- interruption de grossesse, rupture de grossesse extra-utérine, accouchement par forceps ou par césarienne, extraction manuelle du placenta, manipulations sur le bébé afin d’accélérer l’accouchement ;
- morsure de vipère, d’insecte ou d’autres animaux;
- transplantation d’un organe ;
- mycoses favorisées par les plaies ou les crevasses sur la peau qui guérissent mal;
- contact direct des plaies avec des produits animaux.
En réaction, l’organisme produit des anticorps, mais cela n’est pas systématique.
Chaque organisme réagit différemment aux antigènes. La réponse de certains organismes est violente, et ils produisent une grande quantité d’anticorps immuns. D’autres n’en produisent pas ou peu, même après plusieurs contacts avec les antigènes en question. Mais en général, l’introduction répétée d’antigène dans le sang favorise l’augmentation du taux d’anticorps et la sensibilité de l’organisme à cet antigène.
L’agglutination et l’affinité des anticorps
La transfusion de sang doit être précédée par la recherche d’anticorps et d’antigènes car le sang du donneur doit être compatible avec celui du receveur. Le sang compatible contient les antigènes identiques à ceux du receveur mais, en pratique, cette condition n’est pas réalisable car un tel sang n’ont que les jumeaux monozygotes. Cette une raison pour laquelle on utilise un sang semblable, donc un sang du même groupe sanguin du système ABO et Rhésus, c’est-à-dire, un sang aux antigènes bien identifiés.
Une transfusion idéale est celle où le de sang du malade est utilisé. Ce sang doit être récolté et conservé dans des conditions convenables, afin de l’utiliser uniquement pour ce malade en cas de besoin.
En cas d’incompatibilité de sang, les antigènes A se lient aux anticorps naturels et immuns anti-A, les antigènes B se lient aux anticorps naturels et immuns anti-B, et les antigènes D se lient aux anticorps immuns anti-D (les anticorps naturels anti-D n’existent pas). Les autres antigènes se lient aux anticorps spécifiques.
Un anticorps de classe IgG possède deux „bras” pour se lier aux autres molécules, et il peut se lier avec deux érythrocytes étrangers ( si des charges électriques sont faibles).
L’anticorps de classe IgM, malgré les dix „bras” qu’il possède, ne peut se lier qu’à deux-trois érythrocytes dont les dimensions sont 200 fois plus grandes.
Ce rassemblement d’anticorps et d’antigènes est appelé agglutination. Les forces qui lient ensemble ces molécules sont appelées forces de van der Waals et de Coulomb. Ces forces dépendent et varient selon :
- Le nombre d’anticorps : plus ils sont nombreux et plus les forces d’attraction augmentent ;
- Le potentiel zêta : les érythrocytes de haut potentiel repoussent plus fort et la capacité de liaison baisse ;
- Les classes d’anticorps : des anticorps de grandes dimensions (IgM) ont plus grande capacité de liaison avec d’ autres molécules que de petits anticorps (IgG) ;
- L’affinité des anticorps : si les sites de liaisons des anticorps correspondent par leur forme aux sites de liaisons des antigènes, leurs surfaces mises en contact sont grandes et les forces d’attraction les sont aussi. Si les structures des deux molécules sont différentes au niveau des sites de liaison, les forces qui les lient sont faibles.
En cas de faible agglutination, des érythrocytes ne peuvent être détectés qu’au microscope car dans ce cas, les groupes d’érythrocytes sont de petite taille (deux-trois érythrocytes). Une forte agglutination donne des gros amas visible à l’œil nu.
Le système capillaire
Les capillaires dans tout l’organisme ont au total environ 150 000 kilomètres, dont plusieurs centaines de kilomètres dans le cerveau, et la vitesse du sang qui s’écoule est de 0,5 millimètre par seconde environ.
En cas d’hémagglutination, les amas bouchent les capillaires en bloquant la circulation de sang. Plus le nombre de capillaire obstrués est grand et plus l’irrigation sanguine est mise à mal, ce qui provoque une mauvaise alimentation des cellules en oxygène et en nutriments nécessaires pour leur fonctionnement. Une quantité importante de capillaires bouchés peut avoir des conséquences fatales.
Le cerveau, qui constitue 2% du poids de l’organisme humain, utilise 22% d’oxygène fourni par la respiration. Le cerveau doit être approvisionné en permanence, et une interruption d’apport d’oxygène pour une durée de plus de quelques minutes crée des dommages irréversibles [5, 7, 8]. Selon la localisation de l’obstruction et l’étendue de la zone concernée par le manque d’oxygène, on peut observer des signes suivants :
– la somnolence,
– les nausées, les vomissements,
– l’apathie,
– les hallucinations,
– des troubles de la vue et du systèmes nerveux,
– des troubles du système circulatoire, digestif et respiratoire.
D’autres symptômes peuvent survenir, tels que : une hausse de température, des douleurs lombaires, une sensation d’une gorge serrée, un engourdissement des membres, des malaises.Le manque d’oxygène prolongé se termine par le décès.
Notons au passage que les symptômes énumérés ci-dessus ressemblent aux symptômes d’une transfusion mal tolérée ou aux symptômes de toxémie gravidique au cours de la grossesse.
Dans les poumons, le sang coule dans un dense réseau capillaire constitué de plus petits vaisseaux sanguins aux parois très fines, de 7 000 nm de diamètre environ et de longueur de 400 000 nm ( 0,4 mm) chacun. Chez un adulte, la longueur totale de tous les vaisseaux dans les poumons est d’environ 5 000 kilomètres. Les fines parois des capillaires laissent passer des molécules de masse allant jusqu’à 1 kD. La surface totale des parois des capillaire est impressionnante : de 90 à 120 m² environ.
La quantité d’air introduit dans les poumons d’un individu adulte pendant 24 heures est de 15 kilogrammes dont 3 kilogrammes passent dans le sang après avoir traversé les capillaires.
Il faut avoir cela à l’esprit et même si nous sommes habitués à sous-estimer les molécules d’air car elles sont invisibles, le poids de 3 kilogrammes est très parlant. Ce volume d’air peut contenir de grandes quantités de substances nuisibles pour un organisme donné.
Sur le même thème
Les haptènes présents dans l’air expiré par un individu peuvent pénétrer dans les poumons et le sang d’un autre individu, ce qui risque de provoquer diverses réactions indésirables.
Des recherches plus avancées ont permis de découvrir que dans les liquides corporels humains existent les antigènes du système Lewis : LeA et LeB. Ce système permet de déterminer le statut de sécréteur ou de non-sécréteur d’un individu.
Les liquides corporels (sueur, salive, lait maternel, urine, larmes…) de la majorité des individus contiennent de petites molécules solubles dans l’eau. Et parmi eux il y a les antigènes de groupes sanguins, autrement dit les haptènes, les antigènes Lewis et d’autres encore.
Êtes-vous sécréteur ? Il est possible de connaître son propre statut de sécrétion grâce à une simple analyse de sang. Cette analyse nous apprend sur la capacité de notre organisme de faire face aux maladies.
Il est impératif de tenir compte des différences entre les sous-groupes sanguins car le fait de les distinguer lors des actes médicaux permet aux patients d’éviter de nombreux effets secondaires.
Les haptènes de groupes sanguins jouent un rôle important dans l’apparition des maladies, même celles d’étiologie inconnue.